
The human brain is widely viewed as one of the most complex and remarkable known objects. Indeed, after considering its intricate structure and vast functionality, it is easy to admire the beauty of the brain. But, to truly appreciate it, one must consider its past and attempt to understand the context through which human neurophysiology evolved.
As is the case with all biological structures, the brain evolved and developed according to its capacity to increase reproductive fitness. In this view, the purpose of the brain is to successfully transmit genetic information into future generations. Herein lies the curious wonder of the brain: those qualities of cognition, from motor function and sensation to human intelligence and logic, have developed through the principle of reproductive superiority.
This article addresses the processes that likely produced human intelligence. Here, ‘intelligence’ describes the highly refined capacity for logical reasoning, symbolic language, and self-awareness.
The evolution of intelligence is largely considered the consequence of two developments: encephalization, or increased brain size, and structure. Today, researchers are exploring the interaction of the two to better understand how each affects cognition, which evolved first, and why.
Research Methodologies
Before discussing human brain evolution, it is important to consider the difficulties inherent to such research. Because no ancient brain exists for direct examination, scientists must use inferential methodologies such as paleontological fossil record studies and living-species comparisons. Both techniques provide valuable data but are subject to limitations that make such evolutionary research difficult1.
For example, a fossilized skull can provide an indication of brain size as well as some information regarding cortical structure1. By considering the collection of discovered human fossils, researchers can accurately track the general tendency of expanding cranial volume through evolutionary time.
While this increase does correlate with intelligence, it fails to explain processes at the neuron level. In this way, cognition is inferred from external benchmarks, without completely considering the cellular and molecular processes, which give rise to intelligence [2]. Such conclusions are like predictions of automobile speed capacity that are based on vehicle size and shape – without considering the engine.
Furthermore, living-species comparisons require that species are similar enough that such appraisals are valid, but different enough that findings are detectable and meaningful. Yet, these comparisons tend to provide stronger evidence for cellular explanations of intelligence. In recent years, the advances in genetics research have provided additional support for evidence generated through this line of research.
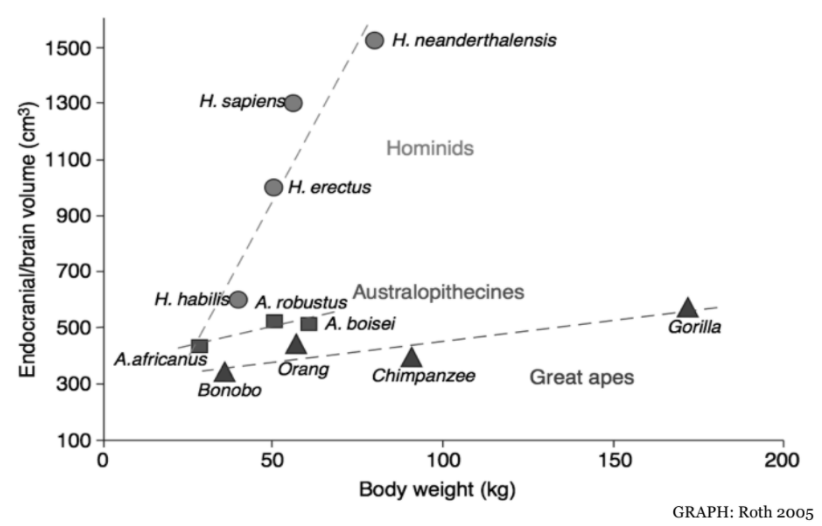
Brain Size
When considering neuroanatomical differences between various species and human ancestors, an obvious dissimilarity lies in brain size (Fig. 1). From a gross anatomical perspective the logic flows easily: more brain tissue, increased processing, higher intelligence.
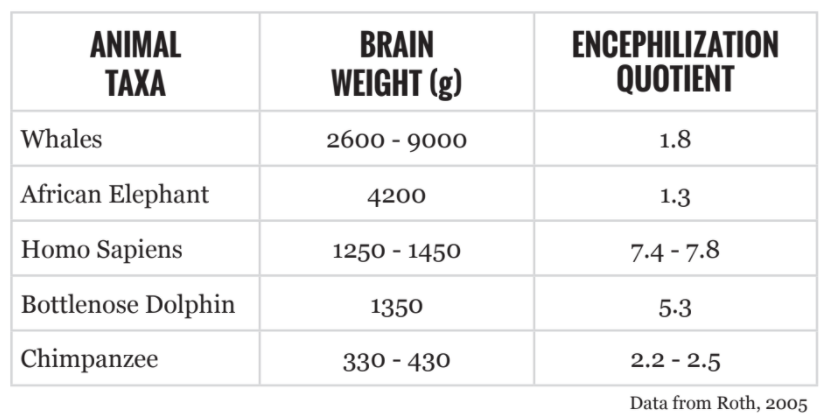
Problematically, there are many instances of larger brains that do not correspond to higher intelligence. Whales, elephants, and some dolphins surpass human brain mass (Table 1).
Even within the Homo genus, there are interesting deviations from the hypothesis that size leads to intelligence. For example, the H. neanderthalensis brain was nearly 200 cm3 larger than modern H. sapiens+. Another such divergence, H. floresiensis, will be considered in greater detail below.
In attempts to compare raw brain mass across species, researchers use a measurement known as the Encephalization Quotient (EQ=Ea/Ee), which considers the ratio of actual brain mass (Ea) to expected brain mass (Ee) for animals of a given size [2]. According to EQ measurements, the largest brains are found in Homo sapiens [2] (Table 1). In other words, humans have brains roughly 7-8 times larger than would be otherwise expected given the species’ physical size.
There are, nonetheless, problems with EQ calculations. Because a sterotyped brain mass is required(Ee), EQ is subject to variation based on categorization. For example, mammal (including H. sapiens) EQ is calcualted using a expected brain mass (Ee) determined by a cat [2]. Because of this EQ can be an unreliable predictor of intelligence.
For instance, some New World capuchin monkeys have higher EQs than chimpanzees and gorillas. Yet, the latter are considered more intelligent [2].
While brain size appears to account for some of human intelligence, there are problems with a model of neurological evolution that regards brain growth as the only determinant of human cognition. First, brain function is not strictly intelligence-oriented; second, metabolic restrictions of brain size may have prohibited such evolution; and third, maternal pelvic size limits cranial volume.
Brain Function Beyond Intelligence
One explanation for the tendency of brain matter to increase with body size is the additional neural function required by larger organisms. Unsurprisingly, as body size increases, the number of motor units, the neuron and skeletal muscles it innervates, increases as well3++.
This rather simple example elucidates an underlying flaw with the notion that brain mass increases necessarily lead to higher intelligence. Due to the multi-functionality of the brain, one cannot accurately assume that brain mass increase inevitably signals a proportional augmentation in intelligence.
In the case of larger animals, the brain is larger partially due to the additional processing required to coordinate movement with increased muscle mass.
Additionally, organisms with highly specialized sensation (e.g. dolphins and dogs) allocate greater neural resources to the operation of these abilities [3]. Whether an intelligence advantage is associated with encephalization depends on which neural structures experience growth.
Simply stated, the brain is too varied to make specific inferences from general measurements.
Metabolic Restrictions
In order to maintain the nutritional requirements associated with their relatively bulky stature, large primates spend many hours in daily feeding activities [4]. Several studies of extant primates indicate that calorie consumption and body size are largely influenced by diet and feeding time. For these species, the limitations of body and brain development are imposed by their abilities to extract energy resources from their environments [4].
Dr. Herculano-Houzel and Katerina Fonseca-Azevedo, researchers at the Federal University of Rio de Janeiro, determined that for animals that consume a raw foods diet similar to that of the common ancestor to humans and primates, metabolism is a significant physiological limiting factor, which necessitates a trade-off between body mass and number of neurons [4].
The researchers propose that the addition of neurons in large primates comes with a fixed cost of 6 kCal per billion neurons5 per day. Using this cost, additional data regarding extant primate feeding habits, and estimated nutritional information, they calculate that members of the Homo species, in supporting enlarged brains, would consistently require nine hours of feeding per day [4].
How, then, did human ancestors overcome the metabolic restrictions on brain growth without resorting to increased feeding times? According to Harvard anthropologist Dr. Richard Wrangham, they invented cooking [6]. Cooking increases the caloric yield of foods by facilitating chewing, digestion, and absorption4. This hypothesis is supported by the recent research by Dr. Herculano-Houzel and her colleague and suggests that the invention of cooking was a prerequisite to encephalization.
Though this hypothesis is somewhat new and not yet widely accepted, it may be that future studies continue to support this premise. If so, it will be clear that some distinct cognitive benefit, unrelated to brain volume and its associated metabolic cost, distinguished precursors of the human lineage from their primate relatives. This resulted in advances in technologies such as fire manipulation and cooking, which predated the rapid expansion of the Homo brain.
Childbirth Constraints
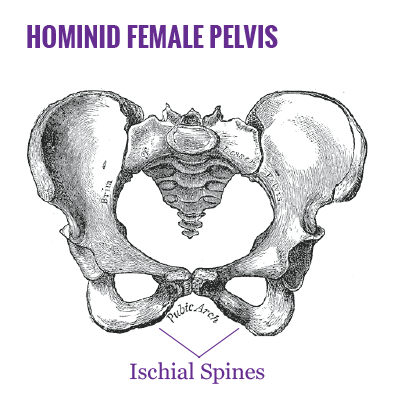
Several hypotheses have proposed explanations for the ultimate cause of hominid bipedal locomotion (efficient long-range terrestrial movement, freeing hands for prolonged food carrying, and possibly tool use). And, while the origin of obligatory uprightness focuses on evolutionary benefits unrelated to intelligence, it may have had a direct effect on the neurological development of the hominid lineage.
With the move to sustained bipedal movement, hominid skeletal structures, particularly the pelvis, underwent mechanical modifications to improve upright balance and to effectively support body weight7. Among these changes, the ischial spines moved toward the midline, restricting the pelvis and reducing the size of the birth canal.
Fossil evidence suggests that bipedalism predates encephalization by at least four million years [7]. Thus, upright locomotion would have imposed an upper limit on cranial volume.
The literature showing the effects of bipedal locomotion on improved intelligence is scarce. For this reason, one is left to ponder the significance of this limit on the evolution of cognition.
It is resonable to assume that as hominid skulls approached this theoretical upper limit, increased intelligence and cognition was unrelated to encephalization. Therefore another mechanism, such as optimization of neural networks, increased synaptic connectivity, or greater cortical surface area, would be required for increased intelligence. Essentially, the existence of a maximum physical volume may have provided sufficient pressure to encourage cognitive gains through efficiency of processing, not increased use of space.
Homo Floresiensis
If encephalization is not the sole or primary catalyst of the forward march of human intelligence, one should expect to find evidence supporting other hypotheses. Indeed, there are many important findings related to genetics, molecular neurobiology, and neuroanatomy, which do offer such examples.
For example, In 2003 a team of archeologists working at an excavation site, known as Liang Bua, on the island Flores, Indonesia unearthed a hominid specimen that was significantly smaller than previously discovered members of the Homo genus. The researchers named their discovery Homo floresiensis+.

Standing at a diminutive 1.09 m (3 ft. 6 in.) and with a cranial volume of a mere 380 cm3 H. floresiensis is exceptionally small [8]. Indeed, its cranial volume is well below the accepted size for the genus Homo and nearly below minimum estimates for Australopithecus [9]. There are several implications of the H. floresiensis find that appear fundamentally incompatible with previously held notions of human evolution10. One such inconsistency appears to be the cognitive capacity of the particularly undersized H. floresiensis brain.
Despite a dramatically smaller brain, researchers have found evidence that the island species exhibited similar cognition as another Late Pleistocene species, H. erectus [10]. Many stone tools and animal remains with evident cut marks were recovered from the dig site; as was evidence of fire usage [8][10].
The technological complexity of H. floresinesis tools led some to initially conclude that they must have been manufactured by modern humans. However, additional examination has indicated that the material and method of manufacture found among the H. floresinesis are distinctly different from later Homo species [10]. Furthermore, these techniques apparently disappear alongside the extinction of H. floresinesis [10].
Despite brains that were 2.5 times smaller than H. erectus, the Flores species were likely much more intelligent. Its brain had unusually enlarged frontal and temporal lobes – areas associated with higher cognition, language, and planning [11].
While the H. floresinesis brain underwent a physical reduction (likely a result of Foster’s rule, also known as the island rule), it maintained higher cognitive function [11]. This indicates that the neurons, lost in the Flores species were likely unrelated to intelligence networks.
While many questions remain unanswered regarding the discovery at Liang Bua, its existence casts uncertainty on a strictly volume-oriented model of brain evolution. Even in terms of EQ, H. floresinesis is only slightly more endowed than modern chimpanzees [9].
Certainly, some mechanism must account for the differences found between the similarly sized H. floresinesis and chimpanzees. If their absolute brain volume and EQ are nearly identical, one must forgo the conclusion that these factors alone explain the greater intelligence of the hominid species. Rather, a more likely explanation is that some distinct neural physiology characterizes H. floresinesis from the extant primate [11][12].
Conclusion
For most of the history of evolutionary brain research, explanations of human intelligence tended to revolve around big-brained hypotheses. This article presents evidence against the notion that species’ brain size is the sole determinant of intelligence. Indeed a rapidly growing series of evidence provides support for cellular and molecular processes that propose possible mechanisms, such as neural organization [12], for human intelligence.
Due to space limitations, this research is not presented here. However, as this is a rapid field of research, future articles, following such findings, will be published here and online at www.greymattersjournal.com/brain-evolution.
Ultimately, the question one must consider when examining the evolution of human neurophysiology is whether intelligence is a consequence of brain volume or a correlated variable.
In reviewing existing evidence, it appears that some cursory intelligence preceded the rapid encephalization of early hominids.