
Most of us are speaking metaphorically when we describe a color as “loud”, a sound as “sharp”, or the feeling of envy as “green”. But for some, these associations are completely literal.
Traditionally, the term “synesthesia” has referred to the peculiar phenomenon in which experiences from one sensory modality can evoke automatic stimulation of a separate one. In synesthesia, the activation of brain regions associated with one sensory modality, the inducer, is involuntarily associated with a second sensory modality, the concurrent.
Current explanations of some common manifestations of synesthesia, such as Visual Motion->Sound and Sound->Color synesthesia rely on the hypothesis that spontaneous cross-activation between associated brain regions due to underlying abnormalities in connectivity is responsible for synesthetic experiences. Recent research supports this hypothesis for synesthesias with inducers and concurrents that are purely sensory, such as color and sound.
There are synesthesias which are not purely sensory, however, in which either the inducer or concurrent represent higher-level features or concepts such as personalities or words which are not properly explained by cross-activation theory.
Dr. Danko Nikolic’s discovery of Swimming Style->Color synesthesia, in which thinking about or performing particular modes of swimming evokes the experience of particular colors prompted him to rechristen this and other higher-level synesthesias as “ideasthesias”.
The existence of ideasthesias has created challenges to understanding synesthesia in general, and has led to the formation of several new explanations for the phenomenon.
What follows is a tour of the different types of synesthesia and ideasthesia, as well as a treatment of the competing explanations for these phenomena, and their respective implications for how perceptual experiences and semantic value are created in the brain.
Experimental Criteria and Objective Verification of Synesthesia
To differentiate between genuine synesthetic experiences and other similar experiences that may be due to associative learning, several criteria have been developed that unite all forms of synesthesia. Among these are the requirement that the concurrent be automatic and involuntarily activated by the inducer in the absence of input which would normally produce it. A genuine synesthetic experience must also be stereotyped, in the way that the inducer must always produce the same concurrent, and their relationship is unaffected by intentional memorization or associative learning.
In general, synesthetic experiences also tend to be simple rather than complex or pictorial [1]. For example, a particular Grapheme->Color synesthete will always see the letter “a” as red. Contrast this to a scenario where viewing a picture of John Lennon would remind a person of the tune for “Imagine” at first, and then “Yesterday” upon second glance. Because the experience is complex, not stereotyped, and is likely the product of associative learning, it is not a genuine synesthetic experience.
Common Forms of Low-Level, Sensory Synesthesia
The form of synesthesia which has been most studied is Grapheme->Color synesthesia, in which letters or numbers are seen in particular colors, as explained in the above example.
Because synesthesia is a subjective experience, early researchers struggled to objectively verify that these experiences occurred and were differentiable from other non-synesthetic experiences. Grapheme->Color synesthesia was among the first of those to be confirmed, due to the ingenious application of several tests invented by Drs. Ramachandran and Hubbard (Fig 1).

Prior to the invention of this test, the Stroop test (Fig. 1A) was thought to be a reliable indicator of synesthesia. However, subsequent studies revealed that non-synesthetes could be trained to associate graphemes and colors in a way that would interfere with their Stroop performance [2].
Tests administered by Ramachandran and Hubbard were designed to differentiate early perceptual levels of processing from later stages that could be contaminated by associative learning (Fig. 1B), and successfully revealed that genuine synesthetic experiences have an observable effect on behavior [3].
Although graphemes can be classified as visual inputs, and therefore appear to be an example of a purely sensory inducer, there is some disagreement about this classification. The identity of graphemes as symbolic units has prompted some to suggest that they have a conceptual component in addition to their sensory input, and that this component may be the inducer [4][5].
Chromesthesia, also called Sound->Color synesthesia, is also relatively common. Synesthetes of this type experience sounds as having particular colors, often with associated geometric patterns or shapes [6]. Particular musical chords are experienced as a combination of these colors and shapes, so chromesthetes can often “see” the contours of a particular piece of music. Because both the inducer and the concurrent are purely sensory, chromesthesia can be considered a low-level synesthesia. For a list of some other common synesthesias, visit www.greymattersjournal.com/synesthesia .
Current Explanations of the Mechanisms of Low-Level Synesthesia
Since only a handful of the known forms of synesthesia have been well characterized, the current explanatory models tend to focus only on these few. Given that the human visual system is relatively well understood, Grapheme->Color synesthesia has been the primary focus of these efforts. It is feasible, however, that explanations for this form of synesthesia can be appropriately generalized to others [3].
Disinhibited Feedback Theory
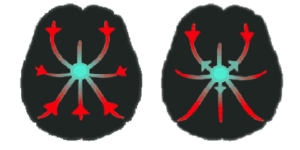
Figure 2A) Disinhibited Feedback Theory
A hypothetical nexus in which sensation (red) may be crossed with perception (green).
The disinhibited feedback model (Fig. 2A) posits the existence of a multisensory nexus, which in non-synesthetes does not send outputs back towards their inputting regions due to properly tuned inhibition. Failure to inhibit the nexus’ output to other sensory areas, however, can hypothetically cause activation of one sensory modality via the nexus when it receives genuine external inputs, resulting in synesthesia.
This theory does not make the assumption that dense interconnectivity exists directly between inducer and concurrent associated regions, but rather that they are connected via a sensory integrating complex, which when malfunctioning can leak activation from one region into another [7].
Cross Activation Theory

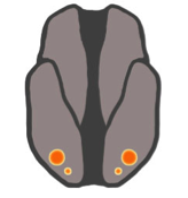
Figure 2B) Cross Activation Theory
Normal connectivity (left) between two sensory regions is not sufficent for cross activation. Synesthete connectivity (right) is much denser.
Cross-activation theory (Fig. 2B) was first proposed as an explanation for Grapheme->Color synesthesia in 2001 by Dr. Vilaynur Ramachandran and Hubbard from the University of California, San Diego. Observations that the visual word form area (VWFA) lies adjacent to the color processing area in V4 suggest that a failure to prune neuronal connectivity between these regions during development results in abnormally dense connectivity between them.
This idea is supported by the fact that synesthesia is known to run in families, and therefore could be a result of genetic differences in pruning of particular regions [7].
Formalization of this model led Ramachandran and Hubbard to make three testable predictions: 1) neurons responsible for the concurrent and the inducer should be heavily interconnected, but not necessarily adjacent, 2) genetic factors are responsible for this interconnectivity, and 3) activation in the neurons that correspond to the inducer should proceed immediately to those of the concurrent [3].
Evidence in support of the cross-activation model’s applicability to Grapheme->Color synesthesia generally takes the form of observed differences in anatomy between synesthetes and non-synesthetes. For example, functional neuroimaging studies have revealed the color selective region of V4 in synesthetes is more active when viewing black graphemes on a white background than viewing non-graphemes in a similar setup, and the heightened activity of V4 was greater in synesthetes than non-synesthetes.
However Ramachandran and Hubbard admit that this result does not point strictly towards cross-activation, and could be consistent with other models such as disinhibited feedback, assuming that the posited multisensory nexus is disinhibited from activating V4.
Anatomical differences in connectivity predicted by the cross-activation model were corroborated by diffusion tensor imaging (DTI, a method for measuring axonal density) studies conducted on 18 Grapheme->Color synesthetes which revealed greater connectivity in the right fusiform gyrus compared to controls [8].
Additionally, the fusiform gyrus and parietal cortex were found via voxel-based morphometry (VBM, a method for measuring cortical volume) to contain a higher volume of grey matter in synesthetes compared to non-synesthetes [9].
As before, these particular anatomical differences support cross-activation theory, but do not rule out other competing theories that posit anatomical differences due to a mechanism other than cross-activation.
The evidence from MEG and EEG studies, however, which allowed researchers to follow the time-course of activation between the regions associated with the inducer and concurrent, favor cross-activation theory as an explanation for Grapheme->Color synesthesia [7].
The third prediction of cross-activation, that the concurrent-associated region will be immediately activated following activity in the inducer-associated region, would predict that V4 would be activated immediately following activity in the VWFA.
The high spatial and temporal resolution of the MEG allowed Hubbard et al. to measure an early activation of V4 111 ms after activation of VWFA in synesthetes that was significantly higher than in controls, a result which is incompatible with disinhibited feedback from a multisensory nexus, which would presumably take longer [7].
This led the authors of this study to conclude that cross-activation is the most feasible model to explain Grapheme->Color synesthesia [7].
Ideasthesia: A Challenge to Cross-Activation
The experiences of Grapheme->Color synesthetes seem fairly well explained within the framework of the cross-activation model.
However the existence of synesthesias that involve higher-level triggers and concurrents, not necessarily adjacent or tied to a particular focal cortical region, suggests that cross-wiring and adjacency of functional brain areas do not describe the mechanism for all types of synesthesia.
This became especially clear after the discovery of Swimming->Color synesthesia4, in which subjects describe the act or imagining of particular swimming styles as possessing particular colors. Unlike Grapheme->Color synesthesia, where the inducer is at least partially sensory, the inducer in Swimming Style->Color synesthesia is purely conceptual.
The discovery of such synesthetic experiences necessitated the differentiation between synesthesias which joined low-level, purely sensory percepts from those involving higher-level percepts that contain semantic value not easily pinned down to one particular region of the brain or modality, also known as Ideasthesias [4].
Dr. Danko Nikolic and his colleagues were among the first to characterize the semantic nature of certain types of synesthesia. Inducers such as swimming style, shape, and time, all of which constitute higher-level percepts, have been shown to evoke concurrents of low-level percepts.
Nikolic was quick to point out that the cross-activation theory and other explanations of synesthesia that rely on basic cross-wiring in the brains of synesthetes are not incompatible with explanations that can account for the role of semantics in synesthesia, but are not able to explain all of the evidence pertaining to synesthetic experiences [4].
For example, several studies have revealed that the context in which certain graphemes are presented will determine the concurrents they elicit: a grapheme which is ambiguously similar both to “S” and “5”, produces different concurrents when it is surrounded by digits versus letters [10].
In his review of this study, Nikolic points out "As the stimulus remained constant, the only variable that changed in these studies was the interpretation of the grapheme, i.e. its understood meaning" [4].
Cross-activation theory, which assumes that synesthesia occurs at a perceptual level, and is not amenable to higher-level influence cannot alone account for the change in concurrent seen in this effect; the impact of semantic, context-dependent cues must therefore be due to a separate mechanism. However Ramachandran and Hubbard have suggested that normal top-down processing of perceptual input that is present in synesthetes and non-synesthetes alike may affect cross-activation, hinting that this may be the separate mechanism [7].
Nikolic also challenges the interpretation of Grapheme->Color synesthetes’ improved performance at serial search tasks such as the one in Fig. 1B. Similar tests, in which synesthetes were asked to locate 2s in a field of 5s, were as difficult for synesthetes as they were for non-synesthetes if the 2s were scattered randomly within the field of 5s. Only when the 2s are arranged in a particular shape for which the subjects are told to search do synesthetes improve relative to controls [4].
The dependence of the synesthetic experience’s ability to confer an advantage suggests that the graphemes themselves do not “pop” in such a grid, and therefore that their coloration relative to other graphemes is dependent on higher-level, conceptual processing, such as concepts of shapes4. In light of the fact that even Grapheme->Color synesthesia may involve high-level inducers, adjacency and cross-wiring of purely perceptual regions of the brain, such as V4 and VWFA, must be complemented with a mechanism that can allow for top-down influence on these regions in order to explain synesthesia.
Additional support for the notion that Grapheme->Color and other synesthesias involve conceptual inducers is the observation that merely thinking about a particular inducer can evoke the concurrent. For example, a grapheme may produce a colored concurrent in synesthetes, regardless of how that particular grapheme is represented (the graphemes 2, II, and “ O O” are all capable of acting as inducers for the same concurrent) [11], and imagining a particular word without actually verbalizing it can evoke concurrent taste in Lexical->Gustatory synesthesia [12].
Theories that Account for Top-Down Influence
Several theories have been put forward to accommodate the influence of top-down input on synesthesia. Of particular interest is the one proposed by Mroczko -Wasowicz and Werning in 2012, which states that mirror-type neurons may act as emulators of sensory experiences, and over activity of these neurons in response to a particular stimulus may result in synesthesia [11].
As an example, they consider the fact that mirror systems have been found that are involved in a wide array of experiences, including motor, sensory, and emotional experiences, i.e. these mirror systems construct a representation of another person’s sensorimotor behavior onto the observer’s own sensory, motor, and emotional percepts.
They liken this to synesthesia in such a way that witnessing another’s behavior is considered the inducer, and the mirror-system mediated sensory, motor and emotional responses are the concurrents. They propose that emulations of the environment through mirror systems would be rooted in neurophysiological pathways, specifically mirror neurons and multi-modal circuits in the intraparietal sulcus and inferior parietal lobule.
Importantly, a particular circuit’s integration of multiple sensory, emotional and motor modalities gives it semantic features (it may relate redness, roundness, a sweet taste, a crisp texture, and the actions of biting and chewing, ultimately contributing to a concept of {apple}), and the circuits responsible for these high-level features may be abnormally active in synesthesia, causing a connected sensory modality to produce a concurrent experience when a particular concept is active [11].
Though still highly theoretical, this and other similar theories present new avenues for experimentation and synesthesia research.
Conclusion
The shift in thinking brought on by the discovery of high-level inducers has forced researchers to reexamine the nature not only of synesthesia, but of the relationship between low-level sensory experiences and high-level conceptual processing. Cross-activation theory’s failure to account for top-down influence on inducers betrays a misunderstanding of this relationship as a coincidental result of the layout of different functional regions, while theories that posit the existence of multi-modal circuits are accommodating of the notion that high-level semantic concepts and their constituent low-level percepts are related non-arbitrarily.
Separately, the existence of synesthesias and ideasthesias serves as evidence in support of qualia and other mental states. The difficulty of finding measurable qualities that correspond to real mental states has in the past led many to favor behavioristic attitudes towards psychology and neurobiology. In light of synesthesia and ideasthesia’s ability to affect task performance and behavior, Behaviorism is less tenable.
Research in synesthesia and ideasthesia will undoubtedly continue to stretch our thinking about the nature of the mind, and will potentially answer more fundamental questions concerning the processes that are responsible for the creation of meaning and our concept of identity.
References
- Cytowic, Richard E. (2002). Synesthesia: A Union of the Senses (2nd edition). Cambridge, Massachusetts: MIT Press.
- Brang, D., & Ramachandran, V. S. (2011).Novel symbols show implicit associations in synesthesia. Manuscript in publication.
- Hubbard, Edward M. “Cross-Activation at Ten.” Journals of Neuropsychology (2011): 152-77. PubMed. Web. 10 Jan. 2013. <http://www.ncbi.nlm.nih.gov/pubmed/21923784>.
- Nikolić, D. (2009) Is synaesthesia actually ideaestesia? An inquiry into the nature of the phenomenon. Proceedings of the Third International Congress on Synaesthesia, Science & Art, Granada, Spain, April 26-29, 2009
- Mroczko-Wasowicz, Aleksandra, and Markus Werning. “Synesthesia, Sensory-Motor Contingency, and Semantic Emulation: How Swimming Style-Color Synesthesia Challenges the Traditional View of Synesthesia.” Frontiers in Psychology 3.3 (2012): n. pag. 22 Aug. 2012. Web. 10 Jan. 2013.
- Cytowic, Richard E; Eagleman, David M (2009). Wednesday is Indigo Blue: Discovering the Brain of Synesthesia (with an afterword by Dmitri Nabokov). Cambridge: MIT Press. pp. 309. ISBN 0-262-01279-9
- Dixon M.J.,Smilek D.,Duffy P.L.,Zanna M.P.,Merikle P.M(2006),“The Role of Meaning inGrapheme-Colour Synaesthesia”, in Cortex,Vol.42,pp.243-252.
- Hubbard, Edward M. “Cross-Activation at Ten.” Journals of Neuropsychology (2011): 152-77. PubMed. Web. 10 Jan. 2013. <http://www.ncbi.nlm.nih.gov/pubmed/21923784>.
- Jancke, L., Beeli, G., Eulig, C., & Hanggi, J. (2009). The neuroanatomy of grapheme-color synesthesia. European Journal of Neuroscience,29 (6), 1287–1293.